Spiral phyllotaxis
Where we use a descriptive model to explain why spirals are
observed in the arrangement of seeds in flowers or scales in
pinecones; where we explain why the number of visible spirals going
in opposite directions are two successive Fibonacci numbers.
Observations in nature
In a daisy or a sunflower, the seeds are not arranged randomly, but
are placed along two families of spirals going in opposite directions
(called the parastichies), and residing approximately on a
disc. The same pattern is observed in pinecones and cactus plants,
although the spirals are stretched along three dimensional surfaces.
Figure 4.1: From left to right: A daisy, a sunflower, a pinecone, a
cactus. First and fourth pictures kindly lend to us by Pau
Atela [3].
Counting the number of visible spirals in the pinecone and the daisy
,(figure 4.2), we respectively distinguish 8/13
(yellow/white) and 21/34 (black/white) spirals in each directions.
These numbers belong to a very curious sequence called the
Fibonacci sequence, 0, 1, 1, 2, 3, 5, 8, 13, 21, Ľ,
where each element is the sum of the previous. It is the aim of this
chapter to explain the reason for these very specific patterns.
Figure 4.2: Presence of spiral patterns in the scales of the pinecone and
the seeds of a daisy.
A descriptive model of Phyllotaxis
Introduction with some examples
In general, the patterns observed in plants is called
phyllotaxis, from the Greek phyllo (leaf) and
taxis (order), and are established already at a very early
stage of the plant's development (in the sunflower, for instance,
these patterns are observed when its flower head is only 2mm in
diameter). At this early stage, one can not talk about seeds or
scales, but small cells known as primordia. Among the
patterns observed, three can be distinguished: whorled, distichous and
spiral, corresponding to different kinds of distribution of botanical
units at a region of the stem (called a node). In whorled
phylotaxis, two or more units grow at a same node; this is observed in
the Archimes Erecta, where three leaves grow at the same
node. In spiral phyllotaxis, only one unit grows at a node, and two
successive units form a constant angle, the divergence angle,
with the tip of the plant shoot (called the apex). Distichous
phyllotaxis is a special case of spiral phyllotaxis where this
divergence angle is 180°, and thereby two successive units are
formed at opposite sides of the apex. This is depicted, in a three
dimensional version, on figure 4.3 (b) (the plant is
a Phalaenopsis).
Figure 4.3: Whorled (a), distichous (b) and spiral (c)
phyllotaxis. All pictures, again, borrowed
from [3].
We will of course study the spiral case exclusively, and refer
to [3] for more information on the other patterns.
Descriptive versus explanatory models
In spiral phyllotaxis, there are two models used to explain the
observed patterns: the descriptive and the
explanatory model. The descriptive one, which we shall
study, uses a simplified, geometric approach, with some assumptions
such as the value of the divergence angle. However, this model does
not care of the dynamical process which the plant, "in reality",
undergo, and this has led to a physical model, the explanatory one,
which describes the interactions between primordia. In this model, the
assumptions are based on hypotheses that Hofmeister [11]
formulated in 1868, as a result of a microscopic study of plant
shoots:
- The newest primordium is formed at the edge of the apex (the
apical meristem) which is usually circular, and moves away
from it radially.
- The period of primordia formation is constant, and is called the
plastochrone.
- The newest primordium is formed where there is most available
space left by the previously formed primordia.
It is the third hypothesis that constitutes the basis for the
explanatory model, since it tells that the coordinates of the newest
primordium are the ones which minimizes a certain potential energy
function. We will not go into more details with this, but complete
studies can be found in [12] and [2].
Modelling spiral phyllotaxis
Notations
It is important to realize that we are not interested in a dynamical
model which describes evolution of the system of primordia in time,
but in a static model, at a given time, where we can analyze the
instantaneous position of the N primordia in the system.
In this system, a primordium is identified by its
index, that is, its position in the sequence the primordia
have been added. Thus, the first primordium added (say, at time t = 0) is denoted p0, the following p1 and so on. When we study the
spiral patterns, we are only interested in the positions of the
primordia, and thus, we assimilate a primordium to a point. And
because of the circular movement of the system, it is convenient to
consider these points by their polar coordinates, i.e. the position of
the i'th primordia is determined by its radius and its angle:
If the phytochrome is denoted P, it is not difficult to figure
out that the i'th primordia is formed at time t = (N - i) P.
Furthermore, assuming that the angle Q between two successive
primordia pi and pi+1 is constant, or, more generally, that
|
Dqn = ( |
®
Opi
|
, |
®
Opi+n
|
) = nQ (2p), n Î N
|
|
and that the first primordia
has angle q0 = 0, pi has angular coordinate qi = iQ at all time t, in accordance with Hofmeister's
first hypothesis of radial movement. Assuming that the velocity of
this movement is strictly positive, we get that every primordia lies
on distinct circles C, the i'th one lying on circle Ci. Its
radius R evolves with time t, i.e. R = R(t) = R((N-i)P). So,
all in all, the position of the primordia in the system, at a given
time, is
|
pi = (i Q, R((N-i)P)), i Î N, i Ł N. |
|
As a consequence, we have to deal with a finite system of primordia,
that is, N < Ą, because otherwise every primordia is infinitely
far from the apex O.
The principle behind the evolution of the system is depicted on figure
4.4 (a).
Figure 4.4: The principle of spiral phyllotaxis. Figure (b) represents
the same model as figure (a), but at a later stage, and with a
smaller plastochrone. Click on the image to see an animation
showing the seeds dispersion in time.
The divergence angle, Q
In most plants, the divergence angle has a very precise value, namely
the Golden angle F = 2 p/ f, where f = (1 +Ö5) / 2 is the Golden Ratio. We shall now give an explanation of this phenomenon,
starting with the following theorem:
Theorem 5:
If Q = [(2 pp)/q], p, q Î Z\{ 0 }, q ł 0, and p/q is an irreducible fraction, then the primordia are
aligned along straight lines distributed uniformly on the pencil [0,2p[.
Proof: Since p/q is irreducible, np/q Î Q\\mathbbZČ{ 0 } for
all n Î N, n < q, so 2pn p / q (2 p), are different
for all n < q. Furthermore, pn Î Z, so it admits a unique
representation pn = mq + n˘, where m Î Z, n˘ Î Z, n˘ < q. Thus, 2 p/ q (mq + n˘) (2p) are different for all
distinct couples (m, n˘). But 2 p/ q (mq + n˘) (2p) = 2 p m + 2pn˘/q (2p) = 2pn˘ (2p), and thereby 2pn˘ (2p) are all diferent for distinct n˘. Since n˘ < q, 2pp n/q (2p) = 2pn˘/ q Î [0,2p[. Therefore, 2pp n / q
divides the interval [0,2p[ into q subintervals of length
2p/q. Moving to the representation in plants, this means that
the primordia are aligned along q straight lines, where the angle
between two neighboring lines is constant, with value 2p/q. This
completes our proof.
Now it should come as no surprise that if Q = 2 pa, where a is rational, the packing of the primordia is
far from optimal, since we obtain a straight lines configuration (such
as the one on figure 4.4 (c), where
a = 1 / 5), leaving large gaps in between. One could then think
that the solution is simply to use an irrational value of a; in
fact, the above mentioned packing will thence not occur, since no two
primordium will ever be aligned with the apex. However, the number
a = f-1 is chosen rather that a = p or a = Ö2, although they are all irrational. But, the latter can be quite
well approximated by rationals, and therefore, the resulting
configuration of the primordium will be very close to the aligned one.
So, not only do we want to avoid aligned configurations, we want to
get as far as possible away from them. In order to obtain that, we
therefore choose the most irrational number, that is, the number which
admits the worst rational approximations.
A trivial but important consequence of theorem
4.2.1 is the following:
Lemma 6:
If Q = [(2 pp)/q], p, q Î Z\{ 0 }, q ł 0, and p/q is an irreducible fraction, then all the primordia
pi+nq, n Î N such that i+nq < N are aligned along
straight lines.
Proof: Simply compute the angle between every q primordia, Dqq = q Q (2p) = 2 pp (2p) = 0, and the result follows.
Now we assume, more generally, that Q = 2pa (2p), where a Î R\{ 0 }. Then we can write Q = 2p(p/q + e), where p / q is a rational approximation
to a, and e = a- p / q. Formula
becomes:
|
Dqq = q Q (2p) = 2pp + 2pq e (2p) = 2 pq e (2p). |
|
In a "sketchy" way, let us join every q primordia
pi+nq, n Î N, n Ł (N-i)/q, starting from one of the
points p0 Ľpq-1. When this has been done for all of the
starting points, we obviously obtain q curves with no common points.
When a has an exact rational approximation, e = 0,
so the angle Dqq between every q primordia is 0.
Thereby we rejoin 4.2.2, where the mentioned curves
are straight lines. But when a is irrational, i.e.
e > 0, the angle between two successive points on the
curve is constant and greater than 0. So, for every constant angular
increment, we move from a point on Ci to a point on
Ci+q.
Try the divergence angle applet, where you
can change the value of the divergence angle in real time and see the
corresponding configuration of the seeds.
Spiral patterns
We will now investigate on the smooth curve passing through the
primordia pi+nq by claiming the following central theorem, which
we might call the fundamental theorem of spiral phyllotaxis:
Theorem 7:
If Q = 2pa, where a is irrational and p/q,
p, q Î Z\{ 0 }, q > 0 is an irreducible rational
approximation to a, and if R(t) is a uniformly increasing
function denoting the radius of Ci at time t = (N-i)P, then
there exists an affine function f : R® R, given by
|
f(q) = P |
ć
č
|
N- |
i + nq
i Q (2p) + nD qq
|
q |
ö
ř
|
, |
|
such that s Ç Ci+qn = pi+nq, "n Î N, n Ł (N-i)/q, where s is a curve given in polar coordinates by
the equation R(f(q)), q Î R.
In other words, this theorem states that when Q = 2pa
where a is irrational, the q curves joining every q
primordia, starting from p0 to pq-1, are all same spirals
(notice that they are not necessarily logarithmic) rotated and scaled
properly.
Proof: By definition, pi+nq Î Ci+nq, so it is sufficient to
prove that pi+nq Î s. As depicted on the picture below, the
point pi+nq has angular coordinate q
= i Q(2p) + n Dqq. From equation , it has radius
coordinate R((N-(i+nq))P). What must be proved is therefore that
s goes through pi+nq, i.e. that R(f(q)) = R((N-(i+nq))P) Ű f(q) = (N-(i+nq))P. By the following computation
|
f(q) = f(i Q(2p) + n Dqq) = (N - (i+nq))P, |
|
we obtain the desired result. For convenience, we will call the
spirals joining every q primordia the "q-spirals".
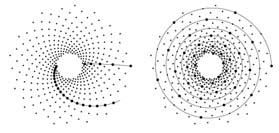 Now we have proved that when the divergence angle is written as
Q = a2p, where a is an irrational number
approximated by the fraction irreducible p/q, we can join every q
primordia by a spiral. If the quality of the approximation is poor, we
will observe very "whorled" spirals. This is due to the fact that
e, and thereby Dqq, is very large. So the angle
between two successive primordia that are joined is large. In
contrast, as the approximation becomes better, the spirals tend to
straight lines, and their number increase since q increases. This is
illustrated on figures 4.5, where the
divergence angle is Q = 2p/f; the radius function R is
the exponential function. Since f is best approximated by the
ratio of two successive numbers in the Fibonacci sequence F, we
have that the 89-spiral (89 Î F) is almost straight, while the
21-spiral (21 Î F) is more whorled, and the 10-spiral (10 Ď F) does not produce a visible pattern at all.
Figure 4.5: Left: 21- and 89-spirals; right: 10-spiral.
Now we have proved that when the divergence angle is written as
Q = a2p, where a is an irrational number
approximated by the fraction irreducible p/q, we can join every q
primordia by a spiral. If the quality of the approximation is poor, we
will observe very "whorled" spirals. This is due to the fact that
e, and thereby Dqq, is very large. So the angle
between two successive primordia that are joined is large. In
contrast, as the approximation becomes better, the spirals tend to
straight lines, and their number increase since q increases. This is
illustrated on figures 4.5, where the
divergence angle is Q = 2p/f; the radius function R is
the exponential function. Since f is best approximated by the
ratio of two successive numbers in the Fibonacci sequence F, we
have that the 89-spiral (89 Î F) is almost straight, while the
21-spiral (21 Î F) is more whorled, and the 10-spiral (10 Ď F) does not produce a visible pattern at all.
Figure 4.5: Left: 21- and 89-spirals; right: 10-spiral.
Visible spirals in plants
We shall now explain, informally, the fact that we actually see
Fibonacci spirals (or for short, F-spirals) in plants, that is,
q-spirals where q Î F. This is not obvious. In fact, in the
above, we have demonstrated that non-Fibonacci spirals are more
whorled than F-spirals. However, what makes us clearly distinguish
some spirals instead of others? What makes us see the 21-spirals of
figure 4.5 instead of the 89-spirals?
Clearly, the quality of the approximation does not affect the
visiblilty of the corresponding spiral. For example, look at the
pattern observed in plants where the seeds have fixed size, such as
the Echinacea purpura below (it will be shown in the next
section that the optimal radius function in this case is the
square-root function). Try to follow one spiral, say from the apex.
You will realize that this becomes more and more difficult as you
approach the boundary, since they will be much more whorled. The same
holds when starting from a visible spiral on the boundary and moving
inwards. This indicates that the number of visible spirals on the
boundary of the flower head changes with the number of primordia N
in the system.
Figure 4.6: The visible F-spirals in Echinacea purpura
are not the same near the apex and near the boundary.
If you look close enough on figure 4.6
(right), you will observe that the spirals are visible when the points
which form it are close to each other. More precisely, they are visible
when the points on the spirals are successively closest neighbors to one
another. So the question is:
why is it always a point pi+n, n Î F, which is the closest
neighbor to pi? In order to answer this, let us consider a
q-spiral, where q Î F. Then the angle between two successive
points on the spiral is given by Dqq = 2 pq e(2p), where e is the error associated with the
approximation of f. Now let us find another fraction p˘/q˘
which approximates f with the same error e, but where
q˘ Ď F. The ratio between two successive numbers of the
Fibonacci sequence being the best rational approximation of
f, it follows that q˘ > q, and thereby that pi+q˘ is
closer to the apex than pi+q, since we move inwards as indices
icrease. But this is not enough to prove that the q-spirals are
more visible than the q˘ ones. We also need to demonstrate that the
angle Dqq is smaller than Dqq˘. We have not
achieved to prove that formally in a general case. However, under the
assuption that e is small enough such that Dqq
and Dqq˘ are smaller that p, we can remove the
modulus from their expression, and hence Dqq < Dqq˘. Combining the smaller angle Dqa between two
successive primordia on the q-spiral together with the smaller
radial distance between these, it follows that pi+q is closer to
pi than pi+q˘.
This informal demonstration can be formalized by studying the
distance between two primordia pi and pi+n,
|
d(pi, pi+n) = |
Ö
|
ri2 + ri+n2 - 2riri+n cosD qn
|
, |
|
by application of the generalized trigonometric formula. Hence, we
seek the values of n Î \mathbbN which minimizes the function. We shall
support the above demonstration by showing two graphs of this
function, evaluated at the discrete points n = 1 Ľ1000 and n = 1 Ľ50, respectively, where the radius function is the
exponential function. We observe that the minimums are found for n
being Fibonacci numbers; from the graph, we deduce that the 21- and
34-spirals are the most visible, while the 13-spiral is only
slightly visible.
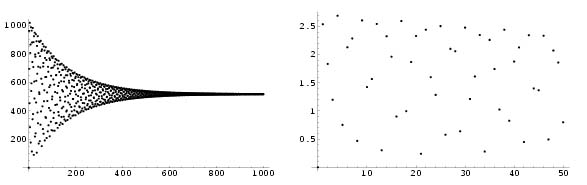 |
<
Figure 4.7: Plot of the distance function between p0 and
pn, n Î 1 Ľ1000 (left) and n Î 1 Ľ50
(right), with R being the exponential function.
Right- versus left-handed spirals
Now that we have explained that the visible spirals are always
F-spirals, it is easy to show that Fn- and Fn+1-spirals are
respectively right- and left-handed, that is, they turn inwards and
outwards in the counterclockwise direction. In fact, the ratio
Fn+1 / Fn approximates the Golden Ratio in an oscillating way,
that is
|
if |
Fn+1
Fn
|
> f, then |
Fn
Fn-1
|
< f. |
|
Thus, f = Fn+1/Fn + e1 where e1 < 0
and f = Fn/Fn-1 + e2 where e1 > 0. As
a consequence, DqFn < 0 while DqFn-1 > 0, yielding opposite directions in the spirals.
Logarithmic spirals in plants?
Figure 4.6 suggests that the seeds in some
plants do not grow once they have been placed. We do not know if this
is true, or if the growth rate is so insignificant that the growth can
not be observed. And in this case, the observed parastichies are, not
surprisingly, no logarithmic spirals, since the phenomenon of growth
does not come to expression here. Concerning the packing of the seeds,
it would be interesting to show which radius function optimizes it. A
straightforward criterion for optimal packing is that the density of
the seeds, say d, is the same everywhere on the flower head (i.e. d = constant). This density is the ratio n/A between a given area
A and the number of seeds n = at, a constant, inside this area.
We are interested in finding the relation between the time t and the
linear dimension, which we call l, that is, a function r such that
l = r(t). We know that the relation between l and A is given by
A = l2 = r2(t). Thus we have d = n/A = at/r2(t). In order to
obtain the seeked constant density, we must have r(t) = b Öt,
b constant. This function can be made equivalent to the radius
function R, and, as a consequence of theorem 4.2.3,
the observed spirals are square-root spiral with polar equation R(t)
linear to Öt.
When the growth rate of the seeds is exponential, in the form
r(t) = a1ebt, it is obvious that the corresponding radius
function is linear to the exponential function, and of the form r(t) = a2ebt. If the radius grows at a different rate, even
exponentially, either the seeds will collide or the will move away
from each other, and thus optimality in the packing clearly does not
hold.
Concluding remarks
It was mentioned in [10] that even for exponential growth,
the corresponding radius function might not be exponential, and with
this function, optimal packings can be obtained. In fact, near the
apex, the seeds need to accomodate immediately after they have been
emmitted, and little time later, the exponential function governs the
radius function completely. So the lack of self-similarity can easily
be neglected in the overall plant pattern.
Another phenomenon often observed in the primordia dispersion
is the change in phyllotaxis type. In fact, often the first primordia
will exhibit distichous phyllotaxis, since there are not enough
primordia to make the pattern stable. For instance, when there is only
one primordia in the system, it is obvious that the next one will be
placed on the opposite side of the apex. The third on will be placed
such that the three primordia-configuration will tend to an isoceles
triangle, and so on. But this pattern will eventually stabilize, and
tends to be of phyllotaxis type spiral.



; ?>/img/seedmovie.gif",320,320,"menubar=no,scrollbars=no,statusbar=no"))
